In beekeeping there are some colony behaviours that we learn about and adapt to, but what is it that makes bees behave in certain ways, some things can be explained others are still a puzzle even to Darwin!
As a beekeeper the evolution of the honeybee can help to explain certain behaviour within the colony. The following attempts to discuss the latest situation.
‘In the Origin of Species, Darwin discussed several challenges that worker insects presented to his theory of natural selection. Complex instincts such as building of combs of hexagonal cells were one problem and were explained by showing plausible intermediate stages. A more serious challenge was posed by the multiple worker castes seen in many ants. How could sterile individuals continue to evolve? Some modern commentaries on Darwin and insect workers seem to be cases of present interests interfering with the interpretation of the past. From a modern perspective, the evolution of a worker caste, and its corollary altruism, are evolutionary puzzles inasmuch as natural selection normally favors greater, not lesser, individual reproduction.’ Darwin’s special difficulty: the evolution of “neuter insects” and current theory. Ratnieks et al
………………………………………………………………………………………………………………………………..
The following is taken mostly from The Buzz about Bees by Jurgen Tautz and explains the kinships of worker bee sisters and half sisters and the drones. It helps to get a feeling about how the bees might behave in certain situations.
William D Hamilton explains how worker bees are more closely related to their sisters than to their own sons which would explain why they are happy to raise their sisters….until we consider the fact that the queen mates with about 13 drones when it becomes apparent that workers are less related to their half sisters than they are to their own sons…Hamilton’s detail is below with my summary at the end.
The unusual situation of kinship in the honeybee colony may be better understood in terms of a concept that has become popular through the work of the English biologist, William D. Hamilton (1936–2000).
The essence of Hamilton’s idea is as follows: particular genes localized at the same place in particular organisms, and which influence the same character, are called alleles. Alleles can occur in different forms, and are the basis for variability in the gene. The alleles are not only passed directly onto the offspring, but copies of these exist also in their siblings and their children, cousins, aunts, uncles, and entire families.
The likelihood of finding the same allele in individuals decreases the more remote the kinship of an individual is to the source. The carrier in which the allele resides is of little consequence for the success a single allele has in spreading as a competing allele in a population.
A behavior in relatives that contributes to rearing the young, for example, can be of advantage to both the supporters and their alleles, even if the carriers have no offspring of their own. Such a sacrifice is then not a disadvantage, if the alleles occur sufficiently frequently in the family.
Kinship selection, a theory developed by John Maynard Smith (1920–2004) and William D. Hamilton, based on the distribution of alleles in groups of related organisms, has clear consequences for the appearance of cooperative, or in extreme cases, “altruistic” behavior in animals.
This theory offers an explanation for single organisms that, like honeybees, have crossed the threshold from “loners” to social beings in the course of their evolution. (Or does it?)
Alleles that are most successful in the branching family network exist “selfishly” at the expense of other alleles. The vision that alleles behave selfishly, and aim only to set as many copies of themselves as possible in the world, has been convincingly explained by Richard Dawkins (1941–), in his book “The selfish gene”.
To an observer, alleles appear as selfishly behaving single elements, exhibiting what could almost be termed a “propagative drive” in honeybees.
Honeybees have, like all other Hymenoptera, and many other insect species that do not form colonies, an unusual mechanism for determining the gender of the adults.
Bees from unfertilized eggs have a single set of chromosomes, the haploid chromosome state.
Bees from fertilized eggs have two sets of chromosomes, the diploid state.
Honeybees possess a single gene for the determination of gender, which can appear in different alleles. An individual that is homozygous for this gene (the alleles are identical), which has to be the case for all haploid individuals (they possess only a single allele), will develop into a male.
An individual that is heterozygous for this gene (all the alleles are different) develops into a female.
A diploid individual homozygous for the sex gene, which very seldom occurs, is a diploid drone, and is usually killed by the workers in the larval stage.
This method of determining the sexes through the number of chromosome sets, or haplo-diploidy, has unusual consequences:
• Males have no fathers, because they come from unfertilized eggs.
It follows that males have no sons, at the most, grandsons.
• Should a male and a female produce daughters, these daughters will share more common alleles than they would with their own children.
Approaching the concept in small steps allows a better understanding of these curious circumstances:
• In 1969, the French bio-mathematician Gustav Malecot (1911–1998) defined genetic kinships as “r”, which is the average probability that a particular allele selected from an individual will also be found in a particular individual to which it is related.
• The value “r” is of biological significance from the point of view of the gene “spender”, because this defines the direction of the gene flow.
• All the alleles of the haploid father will certainly be passed onto each daughter. The probability of occurrence of the father’s alleles in the daughters is 100%, or, expressed differently, r=1.0.
The father will therefore find every one of his alleles again in every daughter.
• The statistical probability that the same alleles of the diploid mother will be found in her daughters lies at 50%, or r=0.5, because a mother contributes exactly half of her alleles to each of her egg cells.
A mother will therefore find, on average, half of her alleles again in a particular daughter.
• The probability that the same alleles will be found in a comparison between full sisters is given by a summary of factors relating to the father and the mother: half of the genome of a female bee comes from the father, and is identical in all full sisters.
Mathematically expressed, this means that 100% of 50% of the sisters’ genes are identical.
The other half of the genome comes from the mother, and has only a 50% probability of being identical in the sisters, because for each gene the mother has one of two possibly different alleles to offer.
In terms of the entire genome, this means 50% of 50%, or 25% are identical.
If one now adds up the values that come from the alleles of the father and the mother, and compares the sisters to one another, one gets 50%+25%=75%, or r=0.75 genetic kinship.
Honeybee sisters therefore share a statistical average of three quarters of their alleles.
In reality, this value swings between 50% of common alleles (only the alleles from the fathers are inherited), and 100% (alleles from both the father and the mother are the same).
Cloned animals are 100% genetically identical; their degree of genetic kinship is r=1.0.
Human children are 50% identical to their parents; here, the degree of genetic kinship amounts to r=0.5.
Honeybees, with their r=0.75, lie between cloned animals and humans.
From this perspective, the best thing that a female bee can do to propagate her genes is to renounce having her own children, and instead help her mother to bring as many sisters into the world as possible.
In order to propagate their alleles, the sterile workers should cooperatively support each other. This is exactly what happens in bee colonies, although the situation is a little more complex.
A queen on her nuptial flight usually pairs with about 13 drones, and their sperm fertilizes the eggs that will later develop into females. The workers in a bee colony all have the same mother, because they all stem from the same queen, but are from many fathers.
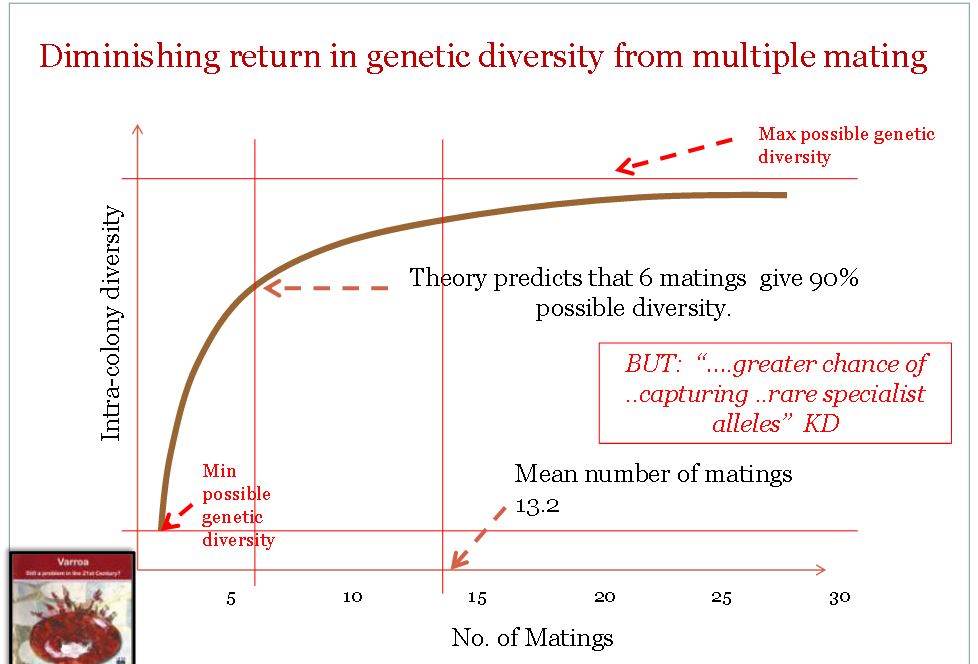
Graph by Glyn Davies of Newton Abbott BKA
Workers that are produced from the sperm of the same drone are full sisters. They are half sisters to those that have different fathers.
Full sisters share more common alleles than do half sisters, so they should support the half sisters less than they do other full sisters.
A complex game of cooperation between the full sisters, and conflict between the full sister groups would be expected if bees supported their closest kin, although an interaction of this kind would depend on them being able to distinguish between full and half sisters.
Bees can determine a great deal about their conspecifics through their sense of smell. The decision of whether or not a bee that wishes to enter the hive belongs to the colony has fundamental importance. This check is undertaken by guard bees at the entrance to the hive, which can smell a newcomer from a distance, and touch her with their antennae when she lands.
Chemo-sensitive sensilla in their antennae enable them to establish whether she belongs to the nest, or is a stranger.
If the odor signals “stranger”, the newcomer will be aggressively chased off. She does, though, have the possibility of being granted entrance if she bribes the guard bees with a drop of nectar.
Conditioning experiments have shown that bees are able to distinguish full sisters from half sisters by the odor of their cuticle, the thin wax layer that covers all insects and protects them from dehydration. Do they use this ability, and if so, when would it be significant in terms of kin selection?
For kin selection, odor identification would be important when new reproductive animals are being reared, because the queens and the drones have a propagative future.
The rearing of a new queen will set the genomic direction for the new colony, and here there is a high potential for conflict between the different groups of full sisters in the nest.
We know virtually nothing about how a colony decides who the new queen will be.
Do subtle conflicts and contests take place between the half sisters that we have not recognized? Do the still generally unknown, but often reported behavior patterns of workers, young queens, and drones on nuptial flights play a role?
Much of this is still a complete puzzle.
An additional area of potential conflict occurs when the workers themselves begin laying eggs.
In European bees, this happens at a rate of 1 in 1,000. Such eggs are unfertilized, and result in haploid drones. In such a colony, therefore, drones can arise that stem from the queen, and have a degree of kinship with her of r=0.5.
Drones that stem from workers have a degree of kinship of r=0.5 with their worker mothers. The degree of kinship between a worker and her brother is r=0.25, and this value is independent of the number of queen pairings, because the mother passes her own genes onto her sons in the unfertilized eggs.
Things get really complicated when one calculates the degree of kinship between a worker and her nephew, the son of one of her sisters. The values that one obtains here are dependent on the number of pairings of the queen on her nuptial flight. If only one pairing took place, the worker would have a kinship of r=0.375 with the sons of her sisters (and in this case, all the workers would be full sisters).
With two possible fathers, the degree of kinship to the nephews sinks to r=0.1875, which is below the kinship of r=0.25 shared with brothers.
If the queen had mated ten times, a kinship of r=0.15 between workers and their nephews results.

Graph by Glyn Davies of Newton Abbott BKA
Purely theoretically then, and considering the usually typical multiple mating of the queen, it would be of genetic advantage to the workers to kill the sons of their sisters, but not their brothers, and on no account their sons, with a kinship of r=0.5.
Workers should therefore suppress nephews that are genetically remote from them, and workers eat the eggs of other workers. They should protect their own eggs, and those of their full sisters, while destroying those of their half sisters, but it is still not clear whether bees can distinguish between the eggs of their full and half sisters.
Workers could also “make sure”, and simply eat up all the eggs that have not come from the queen.
The quantitative determination of the genetic kinship between the members of a bee colony provides the basis for an ambitious theory.
The degree of kinship “r” that is calculated is a statistical average that lies between widely separated extremes.
When a honeybee meets another bee, pupa, larva, or a different egg, she is not confronted with a statistical mean for “r”, but with a concrete single “r”. Can a honeybee determine this value when meeting another individual?
The destruction of haploid drone eggs by the workers shows that they can distinguish between the eggs of the queen and their sisters. The chance distribution of the alleles will, however, lead to situations in which a worker could come across a haploid egg from the queen with which she has nothing genetically in common, or an egg of one of her sisters with which she shares the maximum possible number of alleles.
For the theory to hold, it is not the origin of the egg that determines the action a worker should take, but the nature of the genome.
Just how well honeybees are in reality able to recognize, and use the degrees of kinship still needs to be demonstrated.
In the case of the destruction of worker eggs by workers, there is a simpler explanation: the consumption of eggs could be a purely hygienic precaution.
Very few of the larvae from worker bees molt, and embryonic development either does not start, or the embryo dies. In contrast to determining the degree of genetic similarity, worker bees are faced with the far simpler task of distinguishing dead from living eggs. It is also highly likely that eggs from the queen can be recognized by a protective odor provided by the queen when she lays these. Many questions remain unanswered.
The determination of sex in the form of haplo-diploidy in the Hymenoptera brought about the evolution of superorganisms, and provides an explanation for the change from living as an individual, through living in associations, to sociality and eusociality.
The reality of the presently living superorganisms does not support the theory that kinship alone is the explanation of bee biology. The difficulty of the enormous range of the r-value around the statistical mean has already been mentioned. This becomes even more complicated if the multiple pairing of the queen is taken into consideration when calculating the degrees of kinship.
Hamilton’s quantitative ideas would be valid only if all bees in a colony are from one mother and one father, but because many fathers leave their traces in a bee colony, this does not apply to the bee colonies that we find today.
The workers of a colony are less genetically similar amongst themselves than they would be to their own daughters.
Perhaps we have, in the application of the theory of kinship selection to honeybees, a situation deserving T.H. Huxley’s (1825–1895) remark that “The great tragedy of science is the slaying of a beautiful hypothesis by an ugly fact”. The situation here, though, is not quite as severe. During the passage of evolution, kin selection and haplo-diploidy were needed for the bees, and other hymenopterans, to find their way to their superorganisms.
Hence, when establishing nests, sisters would help one another in raising the young, just as we find today in wasps. But what keeps honeybees still at this level today, if kinship selection is no longer a significant basis?
……………………………………………………………………………………………………………………………………………
A bit more about haploid and diploid:
This diagram is from a lecture given by Gudran Koeniger at the National Honey Show 2014 entitled “Mating Biology of Honeybees. DCA’s – A Natural Strategy To Avoid In-breeding”

In this lecture Gudran also shows results of findings at Drone Congregation Areas, DCA in Austria where drones from 230 colonies were found with over a third of these being the only drone from a particular hive. This demonstrates that the best way to avoid brother and sister mating is to mate outside the hive and at a DCA with drones from many different colonies.
Gudran also mentions the sperm distribution as can be seen in the graph below such that when a queen mates she keeps more sperm from the first drone than the second and so on, hence creating a colony that has an imbalance of groups full sisters. Maybe this imbalance is what influences the choice of new queen or indeed if the colony ever becomes queenless and workers start to lay and raise their own drones.

Some related research notes:
- There is some evidence that bees will selectively confine half-sister queens over super-sister queens, one of the best examples yet of potential genetic control of the final queen. Bee Culture June 2011.
- “Honeybee queens are not reared at random but are preferentially reared from “royal” subfamilies, which have extremely low frequencies in the colony’s worker force but a high frequency in the queens reared.” DNA Analysis of Bees in a Mature Colony (BUT NOW BLUE BLOODED FAMILIES!:- by Robin Moritz, Peter Neumann et al 2005.)
…………………………………………………………………………………………………………………………..
In summary, honeybees still pose a problem for the theory of evolution inasmuch as natural selection normally favors greater, not lesser, individual reproduction. We also see that kinship shows us that sister workers are 75% related which means that they are more related to each other than to their own sons and hence more inclined to raise sisters. However, the queen mates with on average 13 drones and this means that half sisters are less related to another half sister than to their own sons. For honeybees at least then there is still a social harmony puzzle.
Incidentally for bumblebees and wasps kinship can explain nicely how harmony exists as their queen only mates once. But even with bumblebees we see intra colony trouble when the queen switches to drone laying as the workers then prefer to raise their own sons than their brothers. (Goulson)
It is unclear how a queen is selected or indeed the egg or larvae that will become the queen. The logic is that sisters will prefer a queen from their own sister over that of a half sister. Maybe this is the reason that many queens are produced. But the research by Robin Moritz, Peter Neumann et al 2005 would suggest otherwise (see above).
It would be interesting to note how long after the introduction of an unrelated queen to a nuc that the laying of drones ( by the new queen) starts. The obvious logic would be that this would only occur once the majority of the bees in the colony are daughters of the new queen. Yet we see a colony with an unrelated queen that has not mated successfully and hence a drone laying queen has her offspring, her sons reared. Kinship can only be a part of the story of social harmony.
In the scenario above kinship plays no part in the acceptance of a new queen and her sons.
Is it then that the queen pheromone plays a significant part in this harmony?
What we observe is that when a virgin queen emerges she is still immature and the worker bees may recognise her as not being a worker, but they don’t recognise her as a ‘queen’. Over several days the virgin queen starts to mature and becomes ready to mate. She then mates and spends a further few days completing her maturity before starting to lay eggs.
When the workers recognise her as their queen is still unclear, however, it is probably shortly after she has emerged from the pupal cell and become the only queen in the colony. What is also apparent is that her brother drones within the hive treat her as a worker, they don’t try to mate with her. This might be because she doesn’t emit a mating pheromone at this stage. It is highly likely that drones will only mate on the wing and when they detect the queen mating signal (pheromones) as it is known that a brother can and does mate with his sister queen hence the diploid male eggs that the workers reject.
Further study of the drone laying worker scenario might give us a further lead into what it is that bonds the workers of the colony.
It remains that honeybees as individuals don’t follow Darwin’s theory of evolution…
Darwin found a solution that solved these challenging difficulties. The problem depicted would be significantly reduced if one accepted that selection could act not only on the individual, but also on the entire colony. Seen in this light, entire colonies would compete for the largest number of daughter colonies that were reproduced, not of individual bees. Modern evolutionary biology now includes the concept of colony evolution in the term group selection. Just why it is that the individual workers of honeybees, and their relatives, bumblebees, wasps, and ants, do not compete against each other within the colony remains unsettled. Nevertheless, it is precisely this renunciation by the workers of producing their own offspring that the honeybees have used as successful strategy to propagate their own genome.
Observation and experience shows that there are colonies that swarm hardly ever, some that supersede their queen hence never swarming and those that seem to swarm several times a season.
Is it really these latter colonies that are the secret to the evolution of the honeybee?
Beekeepers recognise that swarming is how the colony propagates, but we work with it. To get an excess of honey we need an excess of bees and to this end queen breeders will often look to keep ‘swarminess’ to a minimum. It follows that if all colonies behave in a way to only supersede their queen when she gets old, honeybees would die out.
Where does altruism begin and where does it end?

Reblogged this on Beekeeping365.